Natronomonas pharaonis was isolated from soda lakes where it has to cope with two extreme conditions, high salt concentrations and an alkaline pH of 11. It grows optimally in 3.5 M NaCl and at pH 8.5.
Fig. 1: Lake Zug from Wadi Natrun, Sahara Desert, Egypt (photograph from Antiquity 77, No 296, June 2003). The evaporitic deposits from the lakes of Wadi Natrun have been used by Egypt craftsmen for over 6,000 years (A.J. Shortland, Archaeometry 46, 497-516, 2004). The amount of water in the lakes of Wadi Natrun varies greatly during the year and lakes may fuse or disappear. In the archaeological visit of Wadi Natrun in 2002, lake Gabara, from which Natronomonas has been isolated (Soliman and Truper 1982), could no longer be identified.
Fig. 1: Lake Zug from Wadi Natrun, Sahara Desert, Egypt (photograph from Antiquity 77, No 296, June 2003). The evaporitic deposits from the lakes of Wadi Natrun have been used by Egypt craftsmen for over 6,000 years (A.J. Shortland, Archaeometry 46, 497-516, 2004). The amount of water in the lakes of Wadi Natrun varies greatly during the year and lakes may fuse or disappear. In the archaeological visit of Wadi Natrun in 2002, lake Gabara, from which Natronomonas has been isolated (Soliman and Truper 1982), could no longer be identified.
Two strains were described, strain Gabara (type strain) from lake Gabara in Egypt (DSM 2160) (Soliman and Truper 1982) and strain SP1 from lake Magadi in Kenya (DSM 3395) (Tindall et al. 1984).
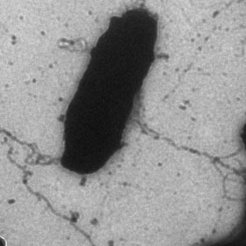
Fig. 2: An electron microscopic image of Natronomonas pharaonis with ca 10.000-fold magnification. Two long flagellar bundles emerge from the rod-shaped cell body. (photo by Christian Klein)
Fig. 2: An electron microscopic image of Natronomonas pharaonis with ca 10.000-fold magnification. Two long flagellar bundles emerge from the rod-shaped cell body. (photo by Christian Klein)
Natronomonas belongs to the order of Halobacteriales, which also includes Halobacterium salinarum , Haloarcula marismortui, and the recently isolated Haloquadratum walsbyi .
Recent publication:
M. Falb , F. Pfeiffer, P. Palm, K. Rodewald, V. Hickmann, J. Tittor, D. Oesterhelt:
Living with two extremes: Conclusions from the genome sequence of Natronomonas pharaonis
Genome Res. 15, 1336-1343 (2005).
The genome of Natronomonas pharaonis
We have sequenced the genome of Natronomonas pharaonis strain Gabara ( DSM 2160 ) ( Falb et al. 2005 ) which consists of the chromosome (2,595,211 bp) and two plasmids (PL131 with 130,989 bp and PL23 with 23,486 bp). A copy of PL23 has integrated into the chromosome and became itself the target for a 13 kb insertion. The chromosome is GC-rich (63.4% GC) featuring four regions of reduced GC content (GC-poor regions I-IV) (see Fig.1).
Fig. 2: Representation of the chromosome of Natronomonas pharaonis. The scaling on top is in Mb. A: Proteins (above line: forward strand, below line: reverse strand) coloured by function category. B: Above line: RNA genes (rRNA operon (red), tRNAs, RNAseP RNA and 7S RNA) . B: Below line: transposase genes, the replication origin (red triangle), and an integrated copy of PL23 with its 13 kb insertion (red bar with white box). C: GC plot and average GC content (blue line). GC-poor regions I-IV are marked. (from Falb et al. 2005)
Fig. 2: Representation of the chromosome of Natronomonas pharaonis. The scaling on top is in Mb. A: Proteins (above line: forward strand, below line: reverse strand) coloured by function category. B: Above line: RNA genes (rRNA operon (red), tRNAs, RNAseP RNA and 7S RNA) . B: Below line: transposase genes, the replication origin (red triangle), and an integrated copy of PL23 with its 13 kb insertion (red bar with white box). C: GC plot and average GC content (blue line). GC-poor regions I-IV are marked. (from Falb et al. 2005)
By rigorous evaluation of the automatic gene finder data, 2843 protein-coding genes were predicted. As automatic gene finder results are highly error-prone for GC-rich genomes, the analysis was based on a number of additional criteria.
Sequence homology between proteins from Natronomonas and the closely related Halobacterium (data from strain R1 ).
Emerging genome-wide proteomic data for Natronomonas .
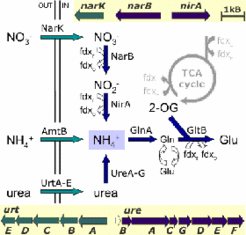
Nitrogen metabolism of Natronomonas pharaonis
Natronomonas grows under highly alkaline conditions in brines of pH around 11 (Soliman and Truper 1982, Tindall et al. 1984). These extreme pH conditions cause reduced levels of ammonia. According to our genome analysis, Natronomonas has three mechanisms that supply ammonia, which is then assimilated into glutamate ( Falb et al. 2005 ).
Direct uptake of ammonia (AmtB)
Uptake of nitrate (NarK) and subsequent reduction to ammonia (NarB and NarT)
Uptake of urea (UrtA-E) which is split by urease (UreA-G) to release ammonia
Fig. 3: Nitrogen metabolism of Natronomonas. The figure shows three transporters (green) for exogenous nitrogen source ammonia (AmtB), nitrate (NarK), and urea (UtrA-E) and enzymes (blue) for reduction of nitrate (NarB+NirA) and hydrolysis of urea (UreA-G). The ammonia is assimilated into the central metabolite glutamate (GlnA+GltB), which is derived from oxoglutarate (2-OG). Redox reactions using ferredoxins (fdx) are indicated. Genes for transporters and metabolizing enzymes are clustered in the genome for both, nitrate and urea conversion. (from Falb et al. 2005)
Fig. 3: Nitrogen metabolism of Natronomonas. The figure shows three transporters (green) for exogenous nitrogen source ammonia (AmtB), nitrate (NarK), and urea (UtrA-E) and enzymes (blue) for reduction of nitrate (NarB+NirA) and hydrolysis of urea (UreA-G). The ammonia is assimilated into the central metabolite glutamate (GlnA+GltB), which is derived from oxoglutarate (2-OG). Redox reactions using ferredoxins (fdx) are indicated. Genes for transporters and metabolizing enzymes are clustered in the genome for both, nitrate and urea conversion. (from Falb et al. 2005)
Ferredoxins appear to be the common proteinaceous electron carrier for functional N-assimilation as well as conversion of 2-oxoacids and aldehydes. Natronomonas contains nine ferredoxin genes which belong to four orthologous groups.
Nutritional self-sufficiency and a simplified synthetic medium
Metabolic enzymes comprise a large number of fatty-acid degradation genes, and the complete set of enzymes involved in biosynthetic pathways leading to amino acids, coenzymes, and prenyl-precursors which are the basic building blocks of archaeal lipids and other components such as menaquinone and retinal. Thus, Natronomonas has a high degree of nutritional self-sufficiency. In agreement with this, we were able to simplify the synthetic medium for this species omitting all amino acids except leucine (see Table 1). Requirement for leucine might be caused by disruption of the gene catalyzing the first step of biosynthesis (leuA_1) in its N-terminal region (Falb et al. 2005).
compound
concentration
compound
concentration
acetate (Na+)
20 mM
pyruvate (Na+)
10 mM
NH4Cl
12 mM
leucine
5 mM
NaCl
3.4 M
KCl
27 mM
Na2CO3
175 mM
MgSO4
1 mM
Na2HPO4
2 mM
NaH2PO4
2 mM
FeSO4
5 µM
CuSO4
4 µM
MnCl3
4 µM
ZnSO4
3 µM
CaCl2
3 µM
pH
9.2
Table 1: A simple synthetic growth medium for Natronomonas pharaonis. (from Falb et al. 2005)
Bioenergetics and the respiratory chain
Natronomonas grows under aerobic conditions and thus respiration is its primary mechanism to gain energy. On the molecular level, respiration is mediated by a series of membrane complexes, the respiratory chain, which - classically - generates a proton gradient that is then used by ATP synthase to produce ATP. However, the alkaline environment results in low extracellular proton concentrations that affect membrane-linked bioenergetics. To overcome this problem, other alkaliphiles use sodium (Na+) instead of protons (H+) as coupling ion between respiratory chain and ATP synthase. However, our experimental data prove that Natronomonas uses protons as coupling ion (see below).
The Natronomonas respiratory chain was reconstructed in silico on the basis of the completely sequenced genome. As this did not allow to identify a complete and functional respiratory chain, we initiated an experimental analysis (see below).
Summarized results of the in silico analysis of the Natronomonas respiratory chain (for further details see Falb et al. 2005). The classical respiratory chain contains five protein complexes (see KEGG). Here, the respiratory chain is described in two sections, the pre- and post-quinone steps:
Pre-quinone steps: Oxidation of NADH and other substrates
Complex I and oxidation of NADH:
A complex I analog is likely as the Natronomonas genome contains a large gene cluster homologous to the nuo cluster of E. coli which encodes type I NADH dehydrogenase. The encoded nuo-like complex is probably functional and produces menaquinole.
However, the nuo-like complex probably does not oxidize NADH as genes for the three subunits nuoEFG forming the NADH acceptor module of E. coli are lacking. Experimental data for other respiratory archaea provide evidence that NADH is not oxidized by type I NADH dehydrogenase.
NADH is probably oxidized by type II NADH dehydrogenase (ndh), which was found in the Natronomonas genome.
Complex II: Succinate dehydrogenase
Complex II of Natronomonas functions as succinate dehydrogenase (Scharf et al. 1997) and produces menaquinole.
Quinone carrier:
The quinone functions as a lipid-soluble two-electron carrier, being reduced to quinole.
In mitochondria and E. coli, ubiquinone is reduced to ubiquinole.
In Natronomonas and Halobacterium, menaquinone is reduced to menaquinole.
Post-quinone steps: Reoxidation of the quinole carrier and terminal oxidation
Alternative I: Quinole reoxidation and terminal oxidation in a single step.
Quinole oxidase:
Quinole oxidases reduce molecular oxygen to water, accepting electrons directly from the lipid-soluble quinole. Quinole oxidase genes have been found in several archaea. The Natronomonas genome encodes three terminal oxidases, but none of these shows sufficient sequence similarity to support the idea that one of them functions as a quinole oxidase.
Alternative II: Quinole reoxidation and terminal oxidation in two steps
Complex III: Quinole reoxidation
In mitochondria, complex III transfers electrons from the lipid soluble two-electron carrier ubiquinole to the one-electron carrier cytochrome-c.
In Halobacterium, genes for a functionally equivalent complex III (petABC), which is likely to transfer electrons from menaquinole to halocyanin, have been identified.
No complex III homologs could be identified in the Natronomonas genome.
One-electron carrier:
Mitochondria and bacteria use the one-electron carrier cytochrome-c, a heme protein with a central iron atom.
The intermediate one-electron carrier in archaea is usually a copper protein (halocyanin in haloarchaea, sulfocyanin in Sulfolobus), a homolog of chloroplast plastocyanin (Scharf and Engelhard 1993).
Complex IV: Terminal oxidation
One of the three terminal oxidases from Natronomonas has been experimentally characterized (Mattar and Engelhard 1997) and is likely to function as a halocyanin oxidase.
The sequence characteristics of the other two terminal oxidases also favor that they function as halocyanin oxidases.
Supercomplexes:
Supercomplexes occur in Sulfolobus and contain complex III and IV components.
The complex III enigma of Natronomonas
In Natronomonas, both post-quinone pathways are incomplete.
No quinole oxidase required for alternative I could be found in the Natronomonas genome.
The alternative II via the intermediate one-electron carrier halocyanin is likely.
However, complex III genes have not been found either in the genome.
We postulate an alternative complex III which awaits identification.
Experimental studies were performed and proved the existence of a functional respiratory chain in Natronomonas. We could further show that protons are used as coupling ion between respiratory chain and ATP synthase, in contrast to other alkaliphiles.
Results of ATP synthesis and proton export (acidification) measurements under different experimental conditions (for further details see Falb et al. 2005):
We demonstrated that protons are ejected upon respiration of Natronomonas.
We measured oxygen-induced increase in ATP levels (oxidative phosphorylation) and oxygen-induced acidification of the extracellular medium.
This proves that protons are ejected during respiration with a subsequent increase in ATP levels.
Both, oxidative phosphorylation and oxygen-induced acidification are sensitive to a protonophore, demonstrating that protons are involved in this process.
We measured light-driven ATP synthesis (photophosphorylation).
We measured a light-induced increase in ATP levels (photophosphorylation) and light-induced alkalinization of the extracellular medium.
Natronomonas contains the light-driven chloride pump halorhodopsin (but does not contain the light-driven proton pump bacteriorhodopsin).
We conclude that chloride is actively pumped into the cell upon illumination. This creates a membrane potential which in turn induces protons (and other cations) to flow into the cell.
This induced flow of protons is responsible for both, ATP synthesis and alkalinization of the medium.
The alkalinization of the medium is not inhibited by addition of a protonophore, showing that the primary process (light-driven pumping of chloride ions) is independent of protons.
In contrast, light-driven ATP synthesis is inhibited by the protonophore, proving that the Natronomonas ATP synthase operates with protons.
We conclude that protons are used as coupling ion between respiratory chain and ATP synthase.
Sustained co-functionality of respiratory chain and ATP synthase requires usage of the same ion.
Analysis of photophosphorylation provides a proof that the ATP synthase of Natronomonas is driven by protons.
Consequently, the respiratory chain of Natronomonas must also generate a proton gradient and thus must contain units generating proton motif force.
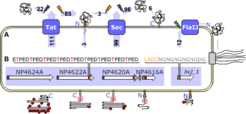
Secretome analysis
Natronomonas faces highly alkaline extracellular conditions, which may have adverse effects on membrane-associated and extracellular proteins. Therefore, we predicted and analyzed the complete secretome. Our data show that a high fraction of the proteins is secreted through the TAT pathway (i.e. in the folded state) as also found for other archaea (Rose et al. 2002, A. Bolhuis 2002). A third of the secreted proteins contain a "lipobox motif" (LAGC), an indicator for covalent lipid attachment (as in halocyanin (Mattar et al. 1994)). This allows anchoring to the membrane to avoid alkaline extraction of the proteins. Covalent lipid anchors are more frequent in haloarchaea than in non-halophilic archaea, the highest fraction being found in Natronomonas.
Interestingly, several lipoproteins have Asn-Gly dipeptide repeats directly following the lipobox. The repeat regions might function as flexible hinges promoting protein-protein interactions.
Fig. 4: Schematic representation of protein secretion, anchoring, and glycoylation. (A) Substrates of the Tat, Sec, and flagellin-specific protein translocation systems with indication of signal peptide cleavage (blue flash signs) and cleavage within the lipobox and subsequent covalent lipid attachment (orange flash sign, orange box). The green flash sign in the flagellin-specific export pathway indicates cleavage by preflagellin peptidase. Numbers indicate how many proteins use a given path. (B) Signal sequence and peptide repeat modules (colored boxes) for a representative gene cluster (white arrows) are presented diagrammatically in models of the cell surface proteins. Indicated are Thr-rich repeats (likely to be O-glycosylated) and Asn-Gly dipeptide repeats. (from Falb et al. 2005)
Fig. 4: Schematic representation of protein secretion, anchoring, and glycoylation. (A) Substrates of the Tat, Sec, and flagellin-specific protein translocation systems with indication of signal peptide cleavage (blue flash signs) and cleavage within the lipobox and subsequent covalent lipid attachment (orange flash sign, orange box). The green flash sign in the flagellin-specific export pathway indicates cleavage by preflagellin peptidase. Numbers indicate how many proteins use a given path. (B) Signal sequence and peptide repeat modules (colored boxes) for a representative gene cluster (white arrows) are presented diagrammatically in models of the cell surface proteins. Indicated are Thr-rich repeats (likely to be O-glycosylated) and Asn-Gly dipeptide repeats. (from Falb et al. 2005)
Signal transduction and motility genes
Natronomonas cells are motile (Soliman and Truper 1982) and actively search for optimal growth conditions. Phototaxis (and chemotaxis) is mediated by a signal transduction cascade similar to that described for Halobacterium. However, there are many differences in the organization of the genes in the genome and the domain architecture of individual components (see Fig. 5).
Cell surface
Thr-rich tetrapeptide repeats varying in length and amino acid composition were detected in nine Natronomonas-specific proteins (see Fig. 4B). Four of the repeat-containing proteins show regional similarity to the cell surface glycoproteins of Halobacterium. Glycosylated Thr-rich tetrapeptide repeats might provide a spacing region that forms a periplasma-like reaction space between cell envelope and membrane.
Fig. 5: The motility and signal transduction clusters of three halophilic archaea. Flagellin (flg) genes are shown in red, genes for flagellin-associated proteins (fla) in blue, chemotaxis (che) genes in green, and genes for additional conserved proteins in yellow. Che genes are separated in two subsets (light and dark green) in Natronomonas and Haloarcula but are arranged into a single cluster in Halobacterium. Similarly, fla genes are separated in two subsets (light and dark blue) in Natronomonas but again are arranged into a single cluster in Halobacterium and Haloarcula. Hatched arrows indicate species-specific duplicated neighbouring genes. Additional conserved genes are dark yellow (present in all three species) or light yellow (present in two species). The MO-ST cluster of Natronomonas is interrupted by a 22 kb insertion. (from Falb et al. 2005)
Fig. 5: The motility and signal transduction clusters of three halophilic archaea. Flagellin (flg) genes are shown in red, genes for flagellin-associated proteins (fla) in blue, chemotaxis (che) genes in green, and genes for additional conserved proteins in yellow. Che genes are separated in two subsets (light and dark green) in Natronomonas and Haloarcula but are arranged into a single cluster in Halobacterium. Similarly, fla genes are separated in two subsets (light and dark blue) in Natronomonas but again are arranged into a single cluster in Halobacterium and Haloarcula. Hatched arrows indicate species-specific duplicated neighbouring genes. Additional conserved genes are dark yellow (present in all three species) or light yellow (present in two species). The MO-ST cluster of Natronomonas is interrupted by a 22 kb insertion. (from Falb et al. 2005)
References
A. Bolhuis: Protein transport in the halophilic archaeon Halobacterium sp. NRC-1: A major role for the twin-arginine translocation pathway? Microbiology 148: 3335-3346 (2002)
M. Falb, F. Pfeiffer, P. Palm, K. Rodewald, V. Hickmann, J. Tittor, D. Oesterhelt: Living with two extremes: Conclusions from the genome sequence of Natronomonas pharaonis Genome Res. 15, 1336-1343 (2005).
C. Klein, C. Garcia-Rizo, B. Bisle, B. Scheffer, H. Zischka, F. Pfeiffer, F. Siedler, D. Oesterhelt: The membrane proteome of Halobacterium salinarum. Proteomics 5, 180-197 (2005)
S. Mattar, M. Engelhard: Cytochrome ba(3) from Natronobacterium pharaonis - An archaeal four-subunit cytochrome-c-type oxidase. Eur. J. Biochem. 250: 332-341 (1997)
S. Mattar, B. Scharf, S. B. H. Kent, K. Rodewald, D. Oesterhelt, M. Engelhard: The Primary Structure of Halocyanin, an Archaeal Blue Copper Protein, Predicts a Lipid Anchor for Membrane Fixation J. Biol Chem. 269, 14933-14945 (1994).
R.W. Rose, T. Bruser, J.C. Kissinger, M. Pohlschroder, : Adaptation of protein secretion to extremely high-salt conditions by extensive use of the twin-arginine translocation pathway. Mol. Microbiol. 45: 943-950 (2002)
B. Scharf, M. Engelhard: Halocyanin, an archaebacterial blue copper protein (type I) from Natronobacterium pharaonis. Biochemistry. 32: 12894-12900 (1993)
B. Scharf, R. Wittenberg, M. Engelhard: Electron transfer proteins from the haloalkaliphilic archaeon Natronobacterium pharaonis: Possible components of the respiratory chain include cytochrome bc and a terminal oxidase cytochrome ba(3). Biochemistry 36: 4471-4479 (1997)
Soliman G.S.H, Truper H.G.: Halobacterium pharaonis sp. nov., a new, extremely haloalkaliphilic archaebacterium with low magnesium requirement. Zbl. Bakt. Hyg., I. Abt. Orig. C 3, 318-329 (1982)
A. Tebbe, C. Klein, B. Bisle, F. Siedler, B. Scheffer, C. Garcia-Rizo, J. Wolfertz, V. Hickmann, F. Pfeiffer, D. Oesterhelt: Analysis of the cytosolic proteome of Halobacterium salinarum and its implication for genome annotation. Proteomics 5, 168-179 (2005)
Tindall B.J., Ross H.N.M., Grant. W.D.: Natronobacterium gen. nov. and Natronococcus gen. nov., two new genera of haloalkaliphilic archaebacteria. System. Appl. Microbiol. 5, 41-57 (1984)
. The evaporitic deposits from the lakes of Wadi Natrun have been used by Egypt craftsmen for over 6,000 years (A.J. Shortland, Archaeometry 46, 497-516, 2004). The amount of water in the lakes of Wadi Natrun varies greatly during the year and lakes may fuse or disappear. In the archaeological visit of Wadi Natrun in 2002, lake Gabara, from which Natronomonas has been isolated (Soliman and Truper 1982), could no longer be identified.")

")
 coloured by function category. B: Above line: RNA genes (rRNA operon (red), tRNAs, RNAseP RNA and 7S RNA) . B: Below line: transposase genes, the replication origin (red triangle), and an integrated copy of PL23 with its 13 kb insertion (red bar with white box). C: GC plot and average GC content (blue line). GC-poor regions I-IV are marked. (from Falb et al. 2005)")
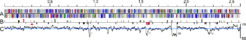
 for exogenous nitrogen source ammonia (AmtB), nitrate (NarK), and urea (UtrA-E) and enzymes (blue) for reduction of nitrate (NarB+NirA) and hydrolysis of urea (UreA-G). The ammonia is assimilated into the central metabolite glutamate (GlnA+GltB), which is derived from oxoglutarate (2-OG). Redox reactions using ferredoxins (fdx) are indicated. Genes for transporters and metabolizing enzymes are clustered in the genome for both, nitrate and urea conversion. (from Falb et al. 2005)")
 Substrates of the Tat, Sec, and flagellin-specific protein translocation systems with indication of signal peptide cleavage (blue flash signs) and cleavage within the lipobox and subsequent covalent lipid attachment (orange flash sign, orange box). The green flash sign in the flagellin-specific export pathway indicates cleavage by preflagellin peptidase. Numbers indicate how many proteins use a given path. (B) Signal sequence and peptide repeat modules (colored boxes) for a representative gene cluster (white arrows) are presented diagrammatically in models of the cell surface proteins. Indicated are Thr-rich repeats (likely to be O-glycosylated) and Asn-Gly dipeptide repeats. (from Falb et al. 2005)")
 genes are shown in red, genes for flagellin-associated proteins (fla) in blue, chemotaxis (che) genes in green, and genes for additional conserved proteins in yellow. Che genes are separated in two subsets (light and dark green) in Natronomonas and Haloarcula but are arranged into a single cluster in Halobacterium. Similarly, fla genes are separated in two subsets (light and dark blue) in Natronomonas but again are arranged into a single cluster in Halobacterium and Haloarcula. Hatched arrows indicate species-specific duplicated neighbouring genes. Additional conserved genes are dark yellow (present in all three species) or light yellow (present in two species). The MO-ST cluster of Natronomonas is interrupted by a 22 kb insertion. (from Falb et al. 2005)")
